

Featured Resources
-
STORM Therapeutics: From Lead to Pre-Candidate Nomination in 18 Months

STORM Therapeutics: From Lead to Pre-Candidate Nomination in 18 Months
Case study
📣From lead to pre-candidate nomination in 18 months. Explore the STORM Therapeutics case study






Discover precise insights into brain neurochemistry with Sygnature Discovery's in vivo microdialysis and cOFM services. With over 20 years of expertise, we design bespoke studies that reveal how compounds modulate neurotransmitter systems in health and disease. Using UHPLC/HPLC with electrochemical detection or mass spectrometry, we deliver robust PK/PD data to support confident CNS decision making.




July 30, 2015

Inobrodib, an exciting, first-in-class oral anti-cancer drug in clinical development by CellCentric, was collaboratively designed, synthesised and supported on its pre-clinical journey by an integrated project team at Sygnature Discovery. Inobrodib is now showing promising results in Phase I and II trials for multiple myeloma and other cancer types.

At Sygnature Discovery, we deliver world-leading drug discovery solutions to accelerate your compound from idea to clinic.

Our leadership team brings diverse experience and insight, driving collaboration and innovation across drug discovery.

Explore careers at Sygnature Discovery and join a global team committed to science, collaboration, and integrity.
Inobrodib, an exciting, first-in-class oral anti-cancer drug in clinical development by CellCentric, was collaboratively designed, synthesised and supported on its pre-clinical journey by an integrated project team at Sygnature Discovery. Inobrodib is now showing promising results in Phase I and II trials for multiple myeloma and other cancer types.
At Sygnature Discovery, we deliver world-leading drug discovery solutions to accelerate your compound from idea to clinic.
Our leadership team brings diverse experience and insight, driving collaboration and innovation across drug discovery.
Explore careers at Sygnature Discovery and join a global team committed to science, collaboration, and integrity.
In this article Riccardo Cavalieri and Steven Harborne discuss the subject and present examples of the current state of play within the field.
Membrane proteins present numerous technical challenges to experimental research. They are expressed at low levels, are often highly dynamic, and can become unstable when removed from the native lipid bilayer. The cell lipid environment, in fact, plays a central role in maintaining their structural integrity and functional dynamics, and its disruption can lead to misfolding, aggregation, or loss of activity. As a result, careful consideration of the membrane mimetic system to be chosen is required from the earliest stages of experimental design.
Choosing a membrane mimetic for membrane protein purification requires balancing biological relevance against experimental tractability. If a membrane protein cannot be successfully extracted from its native membrane and maintained in a soluble, functional state, other factors become largely irrelevant. The choice of membrane mimetic is thus often dictated by the necessity to enable efficient extraction of the target protein, and provide sufficient stability for purification, storage, and subsequent characterisation, rather than by considerations on preserving a “native–like” environment.
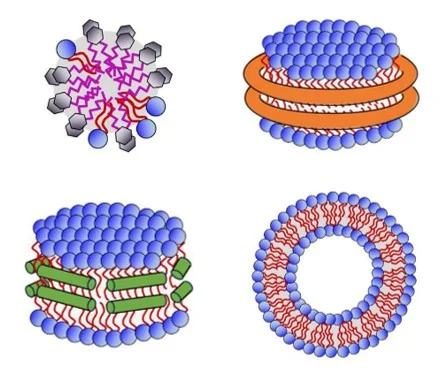
Numerous membrane mimetics with distinct physicochemical properties are available to scientists for the study of membrane proteins. They can be grouped in four broad categories: detergents, polymer lipid particles (PLP) such as styrene maleic acid lipid particles (SMALPs), protein-based nanodiscs, and liposomes.
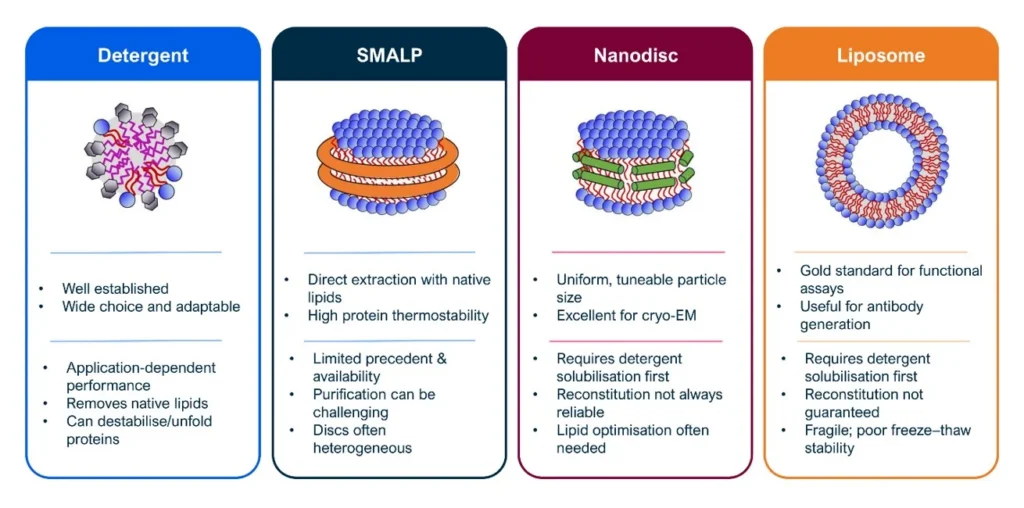
Membrane mimetics are not just passive surfactants that keep the protein in solution, but active physical environments that reshape the conformational behaviour of membrane proteins. Rising evidence from cryo-EM structures, functional assays, biophysical measurements, computational studies, and comparative analyses across detergents, protein nanodiscs, polymers, and liposomes points to the fact that the choice of membrane mimetic fundamentally determines which regions of a protein’s conformational space are accessible or suppressed.
The first point of note is that retention of endogenous lipids does not equate to preservation of native membrane physics. Nanodiscs (polymer or MSP‑based) co‑extract or reconstitute lipids, yet systematically alter their physical properties: lipid packing, lateral pressure profiles, and bilayer elasticity. Whilst polymers that directly extract proteins from the membrane (e.g. SMA, DIBMA, etc) are often thought of as native-like environment, native lipids can coexist with non-native mechanical constraints, leading to conformational biases.
Secondly, stability (especially as measured by thermal shift assays) is often considered a good proxy for correct, physiologically relevant folding and sample quality. While a good stability of the sample is certainly an indicator of tractability for structural studies, recent data on the neurotensin receptor and ABC transporters ABCB10, ABCB25 and LmrA, show that detergents such as LMNG and GDN, as well as small MSP nanodiscs (especially the most used MSP1D1 based), increase stability, homogeneity, and cryo-EM tractability of the protein samples, but do so at the expenses of the conformational ensemble sampled. In both GPCRs and ABC transporters, this stabilisation correlates with reduced ligand efficacy, altered basal activity, and dampened regulatory responses.
Third, recent publications in the literature collectively show that single structures are insufficient descriptors of membrane protein function. High resolution cryo-EM structures frequently capture only the most stabilised state of the protein, and investigating the protein in multiple membrane mimetic environment might be necessary to gain a more complete understanding of its conformational flexibility. An example of this approach has been a paper by Hoffman and colleagues on the ABC transporter MsbA, whose structure was solved in 12 different membrane mimetics (detergents and nanodiscs).
A distinction between structural resolution and functional or biophysical interrogation thus seems to be required when approaching a new membrane protein: no single mimetic optimally serves both objectives, and an exhaustive approach will often need to study the protein of interest in multiple environments.
For single particle cryo-EM applications, success appears to correlate with mimetics that impose strong constraints (high stability and rigidity, low population heterogeneity), such as rigid detergents (LMNG, GDN) or small nanodiscs. These environments are optimal to yield high resolution structures but bias the system towards deep, narrow free energy minima. For proteins with strong lipid dependence (GPCRs, transporters), this often means enrichment of preactivated or artificially stabilised states. Larger nanodiscs (≥18 nm) partially alleviate these effects by reducing lateral pressure, albeit at the cost of increased heterogeneity and potentially lower resolution. Comparative multi environment approaches can provide complementary structural snapshots that collectively represent a broader conformational landscape.
For functional and biophysical assays (e.g. SPR, MST, DSF, etc), the logic is inverted. Mimetics that pre-stabilise basal states or suppress conformational heterogeneity reduce the dynamic range of measurable response. Larger nanodiscs, liposomes, or fast exchange detergents (e.g. DM/DDM) perform better because they preserve the energetic balance required for ligands or substrates to redistribute populations across states. Crucially, assays that measure changes rather than absolute values are the most sensitive to distortions introduced by the choice of membrane mimetic.
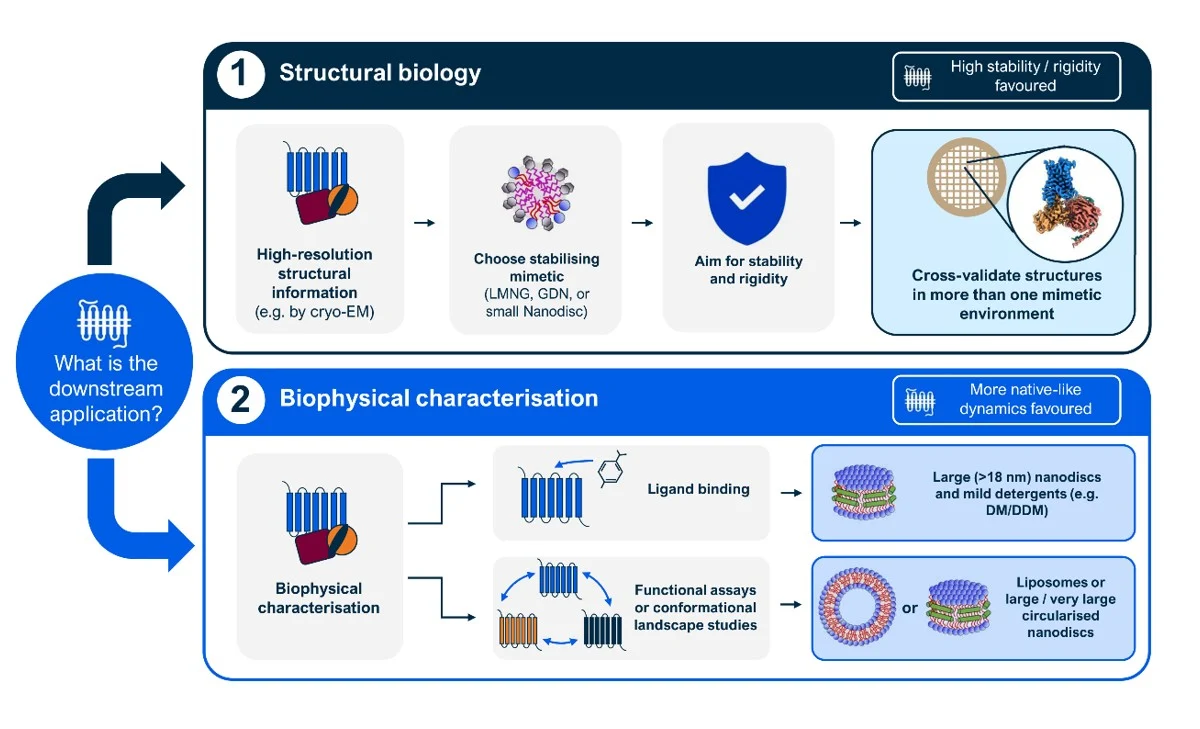
In conclusion, mimetic choice is not simply about preserving compositional similarity to native membranes, or maximising stability of the sample. While the ideal system would preserve the native membrane environment for downstream analyses, it must also allow the protein to be extracted, stabilised, purified, and handled under experimental conditions. If the target protein proves tractable and the choice of membrane mimetics is opened to the scientist, careful consideration is necessary to avoid artificially biasing the protein towards a narrower conformational landscape. Moreover, due attention is to be given to the desired downstream applications for the sample and to selecting approaches that are most appropriate for addressing the research question of interest.
Bibliography





Peak Proteins, NuChem Sciences, and SB Drug Discovery have now fully integrated with Sygnature Discovery.
