

Featured Resources
-
STORM Therapeutics: From Lead to Pre-Candidate Nomination in 18 Months

STORM Therapeutics: From Lead to Pre-Candidate Nomination in 18 Months
Case study










July 30, 2015

Inobrodib, an exciting, first-in-class oral anti-cancer drug in clinical development by CellCentric, was collaboratively designed, synthesised and supported on its pre-clinical journey by an integrated project team at Sygnature Discovery. Inobrodib is now showing promising results in Phase I and II trials for multiple myeloma and other cancer types.

At Sygnature Discovery, we deliver world-leading drug discovery solutions to accelerate your compound from idea to clinic.

Our leadership team brings diverse experience and insight, driving collaboration and innovation across drug discovery.

Explore careers at Sygnature Discovery and join a global team committed to science, collaboration, and integrity.
Inobrodib, an exciting, first-in-class oral anti-cancer drug in clinical development by CellCentric, was collaboratively designed, synthesised and supported on its pre-clinical journey by an integrated project team at Sygnature Discovery. Inobrodib is now showing promising results in Phase I and II trials for multiple myeloma and other cancer types.
At Sygnature Discovery, we deliver world-leading drug discovery solutions to accelerate your compound from idea to clinic.
Our leadership team brings diverse experience and insight, driving collaboration and innovation across drug discovery.
Explore careers at Sygnature Discovery and join a global team committed to science, collaboration, and integrity.
Broad neutral amino acid transporter (B0AT1), encoded by the SLC6A19 gene, is a sodium-dependent neutral amino acid transporter primarily expressed in the intestinal and renal epithelium. B0AT1 operates as a sodium-coupled symporter, co-transporting one sodium ion along with one neutral amino acid across the apical membrane of epithelial cells. For functional expression of B0AT1, both in vivo an in a cell-based assay context, accessory protein TMEM27 co-expression is key to ensure trafficking and stabilisation of confirmation.
B0AT1 plays a crucial role in the absorption of neutral amino acids from the diet in the small intestine and their reabsorption in the kidney1. Mutations in SLC6A10 cause Hatnup disorder, a condition characterized by aminoaciduria and impaired amino acid transport1. Inhibition of B0AT1 has shown potential in normalizing elevated plasma amino acid levels in mouse models of phenylketonuria and urea cycle diorders2. Knockout models of B0AT1 exhibit improved glucose tolerance, elevated FGF21 and GLP-1 levels, and protection against diet-induced obesity3.
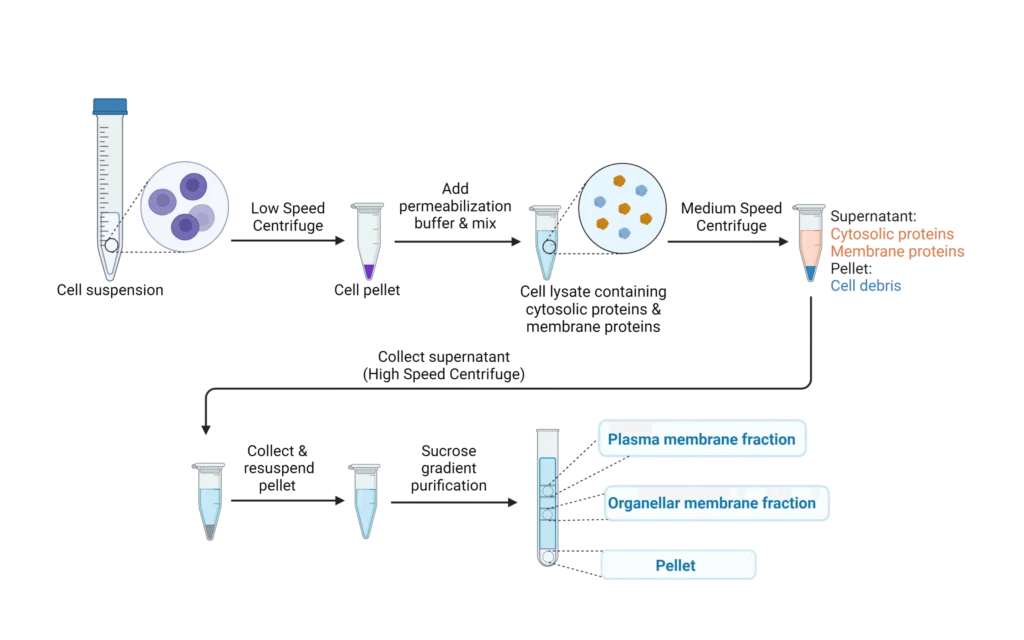
HEK cells expressing B0AT1 and co-transfected with TMEM27 were produced by Sygnature Discovery and harvested in PBS at 80-90% confluency. Untransfected HEK cells served as negative controls. Cells were lysed using nitrogen decompression in a cell disruption buffer. Membranes fractions were isolated through centrifugation and sucrose gradient ultracentrifugation (Figure 1). Protein concentration was assess using Bradford Assay and samples were aliquoted (10 µL) flash frozen and stored at-80°C.
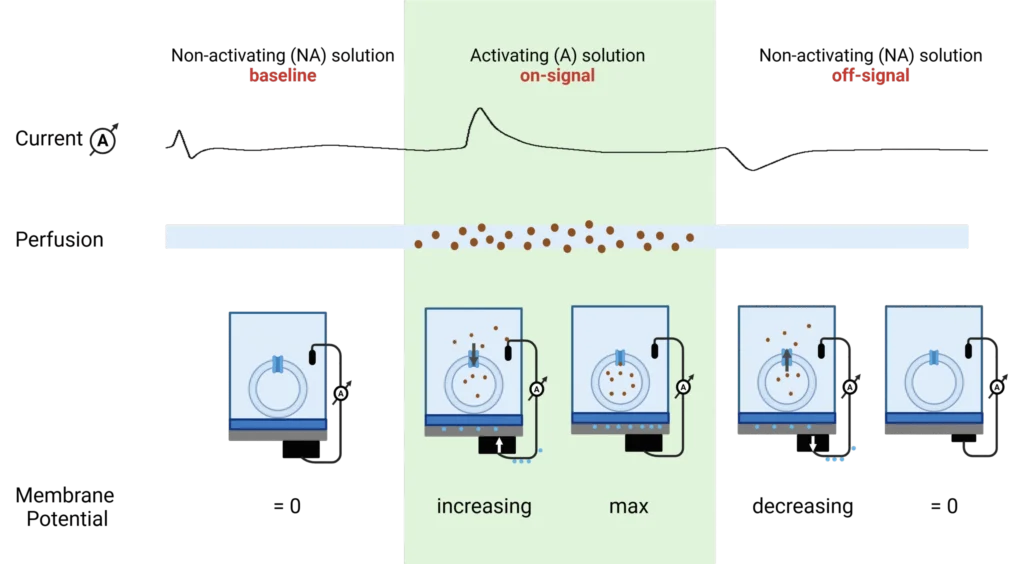
B0AT1 experiments were performed by the exchange of substrate-free (non-activating) buffer for a substrate containing (activating) buffer.
Electrogenic events are measured using SURFE2R N1 and SURFE2R 96SE SSM-based electrophysiology platforms. Charge transloaction is triggered by rapidly perfusing the sensor with a substrate resulting in a gradient across the membrane. The current’s size and shape provided information about speed of transport, coupling ratio or substrate affinity. Data analysis was performed used Data Control 96 V2.3 and GraphPad Prism V10.1.
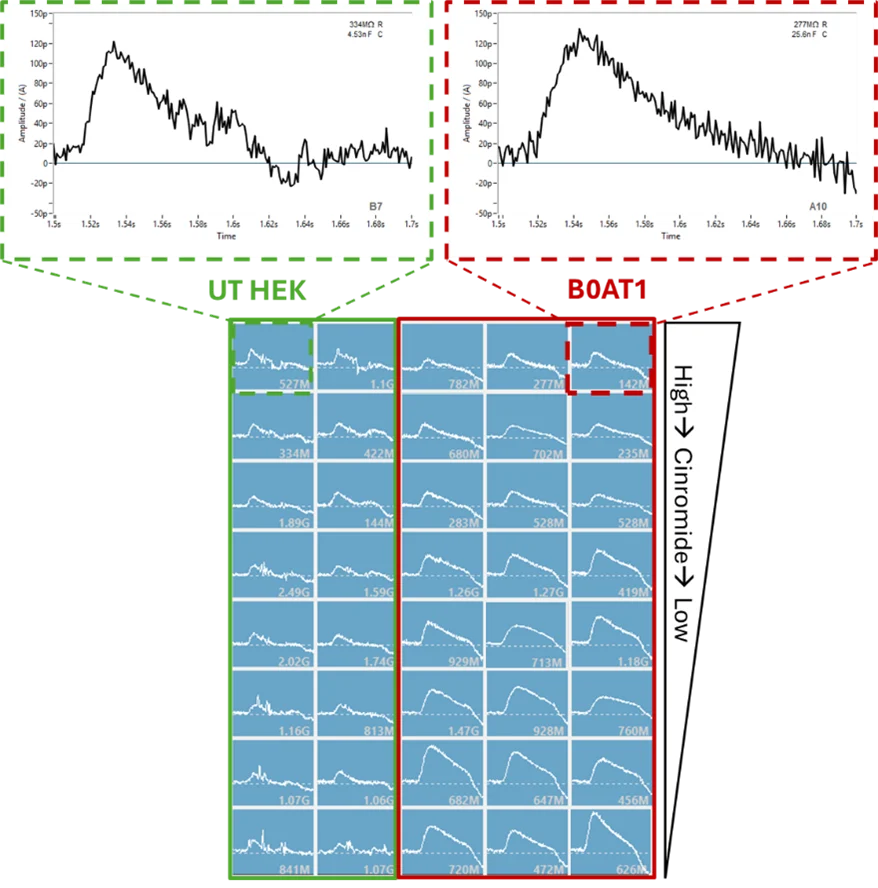
The SURFE2R N1 platform was used for initial validation and SSM-based assay development of B0AT1 (SLC6A19), a sodium-coupled symporter that transports neutral amino acids by harnessing the electrochemical sodium gradient. To confirm sodium dependence, buffers with and without sodium were tested. In the absence of sodium, no transporter activity or net charge movement was observed (Figure 3), confirming the necessity of sodium in the buffer. Negative control experiment using untransfected HEK cells under the same conditions showed only background/endogenous currents, resulting in a 3.65-fold over assay window.
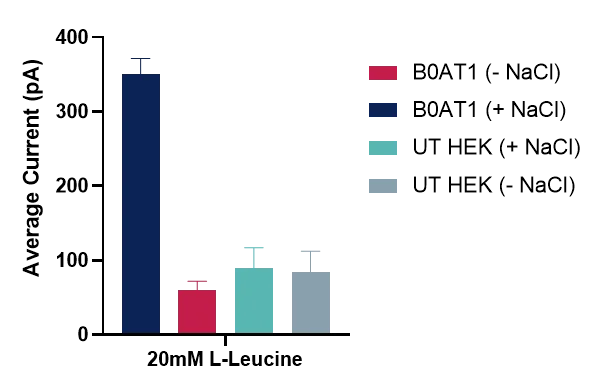
Following establishment of assay conditions and a defined assay window, the dose dependence of transport activity was evaluated. Increasing concentration of L-Leucine were cumulatively applied to sensors containing membranes from B0AT1-expressing or untransfected HEK cells. The averaged responses yielded an EC50 of 1.2 mM and Km of 1.7 mM (Figure 4).
Building on the initial validation of B0AT1 activity with the SURFE2R N1, assays were subsequently transferred to the high-throughput SURFE2R 96SE platform. Conditions established on the N1 were reproducible on the 96SE, and comparison of EC50 values across the two platforms showed agreement within a three-fold window. These results confirmed that both platforms could be used interchangeably with seamless assay translation (Figure 5).
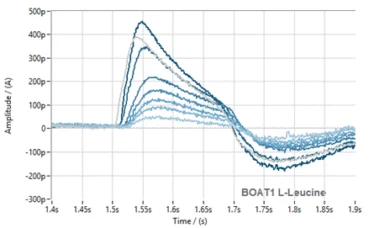
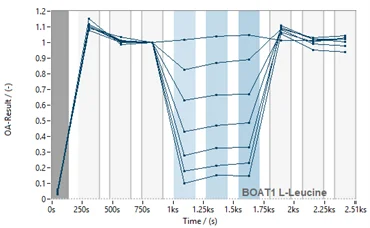
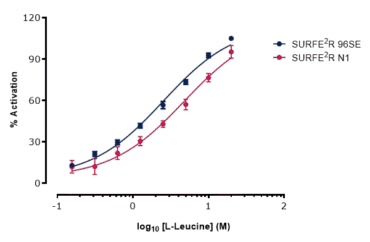
Figure 5. Assay transfer from SURFE2R N1 to SURFE2R 96SE. Normalised current traces (left) and corresponding time course (right) for substrate L-Leucine against B0AT1 on SURFE2R 96SE. Comparison of EC50 values obtained on SURFE2R N1 (red) and SURFE2R 96SE (dark blue) (bottom).
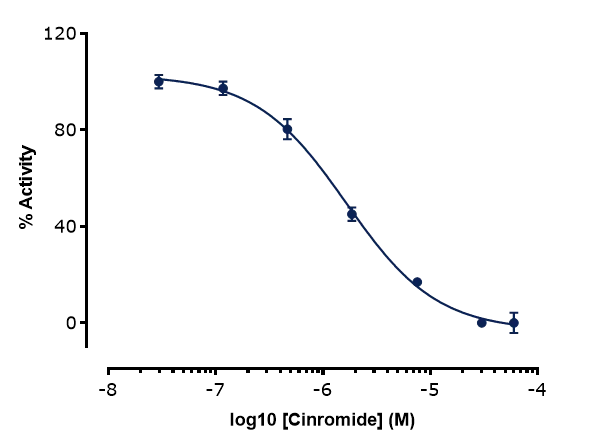
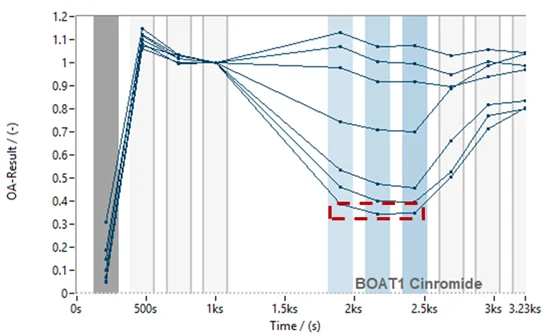
To validate the assay in the 96SE format, a known selective inhibitor of B0AT1, Cinromide (reported IC50 of 0.5 µM in cell-based assays), was profiled. A concentration-dependent inhibition was observed, with a calculated IC50 of 1.6 µM, falling withing a three-fold range of literature values (Figure 6). At the highest concentration tested (60 µM), Cinromide did not appear to fully block B0AT1 signal (Figure 6, top panel, red box). However, closer inspection showed that the residual current was comparable to that observed in untransfected HEK cells (Figure 6, lower panel, red and green boxes). This confirmed that Cinromide fully inhibited B0AT1 activity and that the remaining current reflected endogenous activity from the parental cell line.
Sygnature Discovery’s transporter portfolio spans a wide range of targets supported by both fluorescence-based screening and electrophysiology platforms. Our custom assay development capabilities ensures assays are tailored to fit specific project needs, from hit identification through mechanistic studies and lead optimization. These platforms deliver robust, reproducible data to accelerate transporter-focused discovery programs with confidence.





Peak Proteins has now fully integrated with Sygnature Discovery.
