

Featured Resources
-
STORM Therapeutics: From Lead to Pre-Candidate Nomination in 18 Months

STORM Therapeutics: From Lead to Pre-Candidate Nomination in 18 Months
Case study
📣From lead to pre-candidate nomination in 18 months. Explore the STORM Therapeutics case study






Discover precise insights into brain neurochemistry with Sygnature Discovery's in vivo microdialysis and cOFM services. With over 20 years of expertise, we design bespoke studies that reveal how compounds modulate neurotransmitter systems in health and disease. Using UHPLC/HPLC with electrochemical detection or mass spectrometry, we deliver robust PK/PD data to support confident CNS decision making.




July 30, 2015

Inobrodib, an exciting, first-in-class oral anti-cancer drug in clinical development by CellCentric, was collaboratively designed, synthesised and supported on its pre-clinical journey by an integrated project team at Sygnature Discovery. Inobrodib is now showing promising results in Phase I and II trials for multiple myeloma and other cancer types.

At Sygnature Discovery, we deliver world-leading drug discovery solutions to accelerate your compound from idea to clinic.

Our leadership team brings diverse experience and insight, driving collaboration and innovation across drug discovery.

Explore careers at Sygnature Discovery and join a global team committed to science, collaboration, and integrity.
Inobrodib, an exciting, first-in-class oral anti-cancer drug in clinical development by CellCentric, was collaboratively designed, synthesised and supported on its pre-clinical journey by an integrated project team at Sygnature Discovery. Inobrodib is now showing promising results in Phase I and II trials for multiple myeloma and other cancer types.
At Sygnature Discovery, we deliver world-leading drug discovery solutions to accelerate your compound from idea to clinic.
Our leadership team brings diverse experience and insight, driving collaboration and innovation across drug discovery.
Explore careers at Sygnature Discovery and join a global team committed to science, collaboration, and integrity.
Proteins are remarkably versatile molecules, but some of the most fascinating members of this family go beyond simple amino‑acid chemistry. Many proteins rely on cofactors—non‑protein components that enable or enhance biological activity. Among these, metalloenzymes stand out as a diverse and essential class. Metalloenzymes contain tightly bound metal ions that are integral to their structure or catalytic function. These metal ions can facilitate electron transfer, stabilize charged intermediates, or directly participate in bond formation and cleavage during catalysis, thereby enabling transformations that would be energetically unfavorable or impossible for purely organic enzymes.
Metal coordination in biology is everywhere. Iron in haemoglobin binds oxygen for transport (Terrell J.R., et al. 2018) (Figure 1a-b), magnesium in ATP-dependent enzymes stabilizes phosphate groups (Kwangho N., et al. 2024) (Figure 1c-d), and zinc plays structural and catalytic roles in zinc finger proteins and proteases (Wu, M. et al. 2024) (Figure 1e-f). The precise geometry and coordination chemistry built into metalloenzymes illustrate evolution’s ability to harness metal ions with astounding specificity.
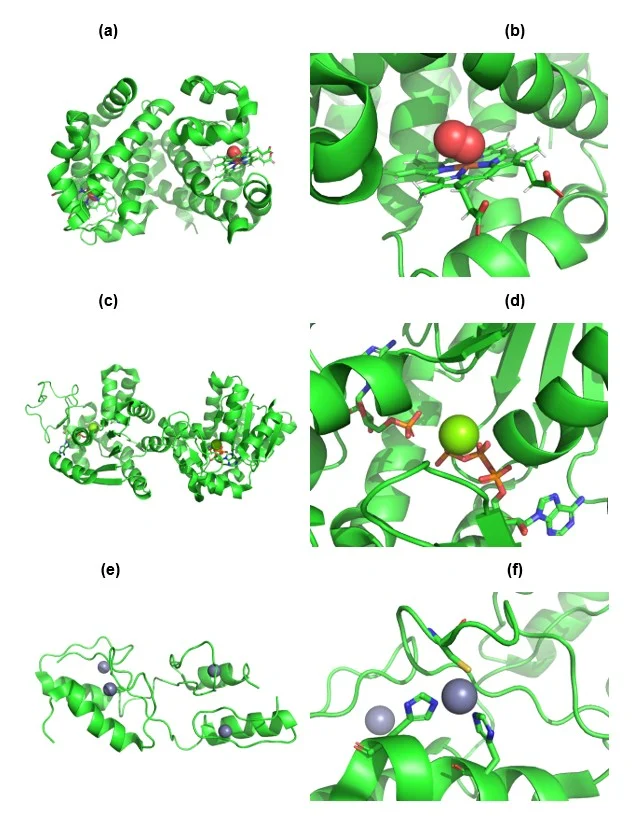
Figure 1. Representative examples of biologically essential metal coordination.
(a–b) Iron coordination in haemoglobin (PDB: 6BB5), illustrating the central Fe²⁺ ion bound within the porphyrin ring and enabling reversible oxygen binding (Terrell et al., 2018). Oxygen shown as spheres. Porphyrin ring shown as sticks. Iron (Fe) atom shown in brown. (c–d) Magnesium-dependent activation of adenylate kinase (PDB: 8RJ6), where Mg²⁺ stabilizes ATP phosphate groups and induces structural reorganization of the active site (Kwangho et al., 2024). Mg ion shown as a sphere. ATP molecule shown as sticks. (e–f) Zinc coordination in GLI1 zinc finger motifs (PDB 7T91), highlighting Zn²⁺‑mediated structural stabilization essential for DNA binding (Wu et al., 2024). Zn ion shown as spheres. Co-ordinating His and Cys residues shown as sticks.
Organisms also make use of more unusual metals. For instance, copper is utilised by the tyrosinase family of metalloenzymes.Tyrosinases are type‑III dicopper metalloenzymes, defined by a binuclear copper active site that is essential for their catalytic function. Each tyrosinase contains two copper ions (CuA and CuB), housed within the central catalytic domain and held in place by six conserved histidine residues—three coordinating each copper ion. This highly conserved histidine-based coordination environment is a structural hallmark of the entire enzyme family (Figure 2) (Pretzler M, 2024).
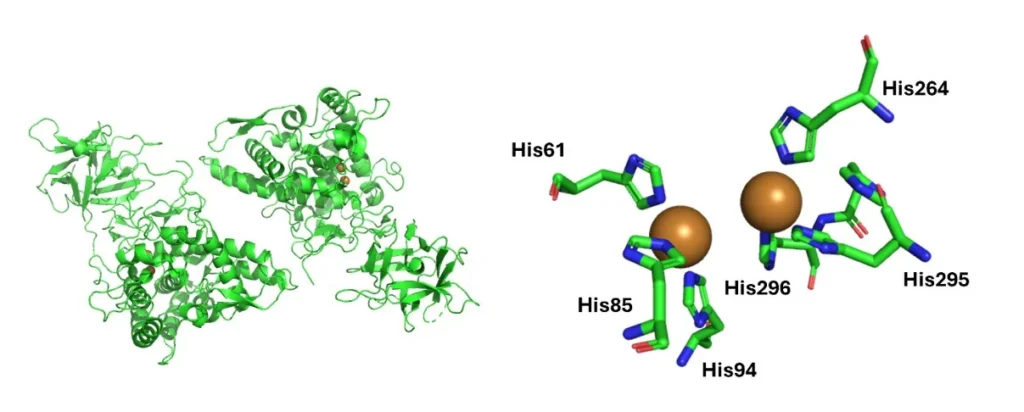
Figure 2. The active site of Agaricus bisporus tyrosinase AbPPO3 (PDB ID: 2Y9W) is shown, highlighting key structural elements that underpin the function of this type‑III dicopper metalloenzyme. Tyrosinases contain two copper ions (CuA and CuB), each coordinated by three highly conserved histidine residues that form the characteristic histidine‑based binuclear copper site essential for catalysis.
The precise geometric arrangement of these copper ions enables tyrosinase to bind and activate molecular oxygen, which is required for both monophenol hydroxylation and diphenol oxidation. The dicopper centre transitions through multiple oxidation states during catalysis, such as the met (Cu²⁺–Cu²⁺) and oxy (Cu²⁺–O₂²⁻–Cu²⁺) forms. This redox flexibility is made possible by the tight histidine coordination, which stabilizes oxygen intermediates and orients substrates within the active site. Mutations of the His-residues or nearby residues which impact Cu binding to tyrosinase, have been known to cause oculocutaneous albinism, specifically OCA1. Tyrosinase produces the precursors of the melanin synthesis pathway, without these precursors the amount of melanin produced is reduce or completely abolished. This ultimately causes a reduction, or absence, of pigment in the individuals’ eyes, hair and skin (Lin, S. et al. 2022) (Kalahroudi, V.G. et al. 2014).
Understanding metalloproteins is one thing—but isolating them in the lab is another. One of the most widely used strategies in modern protein purification relies on metal coordination chemistry, specifically through immobilised metal affinity resins.
Immobilized metal affinity chromatography (IMAC) exploits the interaction between metal ions, such as Ni²⁺ or Cu2+, and histidine residues. By engineering proteins with poly‑histidine tags (typically a His6-tag), we can selectively bind them to nickel-charged resin. Unwanted proteins wash away, while our His-tagged protein elutes cleanly with imidazole. This method is reliable, scalable, and gentle enough to preserve protein structure and activity.
Metals and engineered protein tags are essential tools in protein science because both rely on precise coordination chemistry: natural metal cofactors bind specific amino acids to drive catalysis, stability, and structural organization, while engineered tags exploit similar interactions to purify or immobilise proteins. Metal centres are particularly valuable in high‑resolution biophysical techniques such as X‑ray crystallography and EPR/ESR. Mutating metal‑binding residues (His, Cys, Asp, Glu) helps researchers probe function, as shown in studies linking tyrosinase histidine mutations to OCA1.
In the lab, His₆‑tags enable efficient purification through IMAC, where histidine binds strongly to Ni²⁺ resin, allowing rapid, gentle, and scalable isolation of folded proteins for use in enzyme assays, binding studies, reconstitution experiments, and structural analysis. His‑tags also support controlled immobilization on Ni²⁺–NTA sensor chips in SPR, maintaining consistent orientation for accurate kinetic measurements. However, the placement of tags is critical, as poorly positioned tags can disrupt natural metal coordination, alter folding, or block interaction surfaces, making careful design essential for reliable biochemical and biophysical assays.
Ghodsinejad Kalahroudi, V., Kamalidehghan, B., Arasteh Kani, A., Aryani, O., Tondar, M., Ahmadipour, F., Chung, L. Y., & Houshmand, M. (2014). Two novel tyrosinase (TYR) gene mutations with pathogenic impact on oculocutaneous albinism type 1 (OCA1). PLOS ONE, 9(9), e106656. https://doi.org/10.1371/journal.pone.0106656
Lin, S., Sanchez‑Bretaño, A., Leslie, J. S., Williams, K. B., Lee, H., Thomas, N. S., Callaway, J., Deline, J., Ratnayaka, J. A., Baralle, D., Schmitt, M. A., Norman, C. S., Hammond, S., Harlalka, G. V., Ennis, S., Cross, H. E., Wenger, O., Crosby, A. H., Baple, E. L., & Self, J. E. (2022). Evidence that the Ser192Tyr/Arg402Gln in cis tyrosinase gene haplotype is a disease‑causing allele in oculocutaneous albinism type 1B (OCA1B). npj Genomic Medicine, 7(2). https://doi.org/10.1038/s41525‑021‑00275‑9
Nam, K., et al. (2024). Magnesium‑induced structural reorganization in the active site of adenylate kinase. Science Advances, 10, eado5504. https://doi.org/10.1126/sciadv.ado5504
Pretzler, M., & Rompel, A. (2024). Tyrosinases: A family of copper-containing metalloenzymes. ChemTexts, 10(4), 12. https://doi.org/10.1007/s40828‑024‑00195‑y
Terrell, J. R., Gumpper, R. H., & Luo, M. (2018). Hemoglobin crystals immersed in liquid oxygen reveal diffusion channels. Biochemical and Biophysical Research Communications, 495(2), 1858–1863. https://doi.org/10.1016/j.bbrc.2017.12.038
Wu, M., Jahan, N., Sharp, A., Ullah, A., Augelli‑Szafran, C. E., Zhang, S., & Boohaker, R. J. (2024). Structure characterization of zinc finger motif 1 and 2 of GLI1 DNA‑binding region. International Journal of Molecular Sciences, 25, 13368. https://doi.org/10.3390/ijms252413368




Peak Proteins, NuChem Sciences, and SB Drug Discovery have now fully integrated with Sygnature Discovery.
