

Featured Resources
-
STORM Therapeutics: From Lead to Pre-Candidate Nomination in 18 Months

STORM Therapeutics: From Lead to Pre-Candidate Nomination in 18 Months
Case study
📣From lead to pre-candidate nomination in 18 months. Explore the STORM Therapeutics case study










July 30, 2015

Inobrodib, an exciting, first-in-class oral anti-cancer drug in clinical development by CellCentric, was collaboratively designed, synthesised and supported on its pre-clinical journey by an integrated project team at Sygnature Discovery. Inobrodib is now showing promising results in Phase I and II trials for multiple myeloma and other cancer types.

At Sygnature Discovery, we deliver world-leading drug discovery solutions to accelerate your compound from idea to clinic.

Our leadership team brings diverse experience and insight, driving collaboration and innovation across drug discovery.

Explore careers at Sygnature Discovery and join a global team committed to science, collaboration, and integrity.
Inobrodib, an exciting, first-in-class oral anti-cancer drug in clinical development by CellCentric, was collaboratively designed, synthesised and supported on its pre-clinical journey by an integrated project team at Sygnature Discovery. Inobrodib is now showing promising results in Phase I and II trials for multiple myeloma and other cancer types.
At Sygnature Discovery, we deliver world-leading drug discovery solutions to accelerate your compound from idea to clinic.
Our leadership team brings diverse experience and insight, driving collaboration and innovation across drug discovery.
Explore careers at Sygnature Discovery and join a global team committed to science, collaboration, and integrity.

Indeed, Parkinson’s disease is the second most common neurodegenerative disorder that affects >10 million people worldwide, but unfortunately, to date no disease modifying therapies are available. The economic impact of Parkinson’s disease in just the USA alone is estimated to be around $52 billion which, coupled with the devastating effects of these diseases, means that αSyn is the subject of significant research efforts aiming to develop novel therapeutics, along with methods that will allow an early diagnosis of the conditions.
αSyn is predominantly expressed in the brain [6] and is found at high levels in the presynaptic boutons of axons [7]. It has also been detected in erythroid cells [8] and at low levels in other tissues but, in spite of intense study, the precise physiological roles and function of αSyn still remain unclear (see [4] for a review on the current status of what is known about the function of αSyn). One theory that is gaining support is that αSyn functions to promote membrane curvature, thereby contributing to synaptic trafficking and vesicle budding [9], [10].
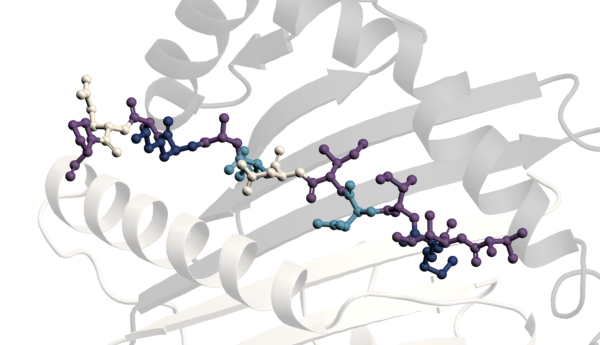
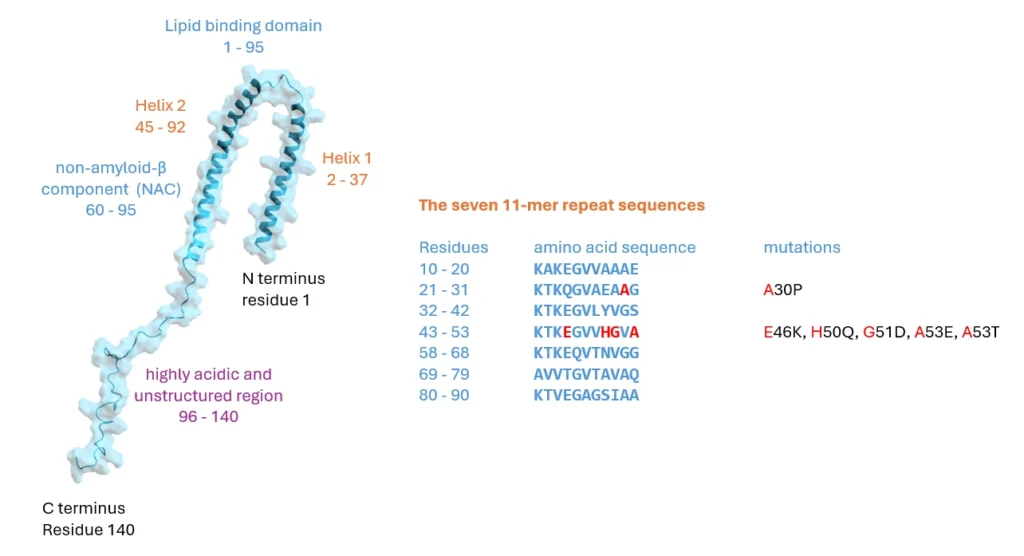
Structurally, αSyn is an intriguing protein. Its N-terminal sequence is divided into seven highly conserved 11-mer repeats with a KTKGEV consensus sequence (residues 1–95), which, similar to apolipoproteins, form an amphipathic alpha-helix with 3 turns, and mediate association of αSyn with lipid membranes. This region also contains the NAC domain (residues 60–95), an area believed to be responsible for αSyn aggregation [2] and sensing of lipid properties [11]. Of particular note is that all identified mutations that are associated with synucleinopathies are located in this region: A30P, A30G, E46K, H50Q, G51D, A53E, A53V and A53T [12]–[20] six of which cluster within eight residues, suggesting that lipid binding or lack thereof may be linked to αSyn pathology. The C-terminus of αSyn (residues 96–140) is highly acidic and largely unstructured [21]–[23] and is the target of various post-translational modifications [24]. The C- terminus has been implicated in modulating αSyns interaction with proteins, metal ions, polycation and polyamine, regulating both its membrane binding and nuclear localisation and protecting αSyn from aggregation (see [3] and references therein). Acetylation of the N-terminal methionine has been shown to be important for oligomerisation of αSyn in diseased states [25].
In Parkinson’s disease, there is mounting evidence that tiny amounts of misfolded αSyn species can spread between cells (perhaps in a prion-like manner) and seed the aggregation of the normal, functional αSyn to form amyloidal plaques [27]. The fibrils of aggregated αSyn comprise a major part of the amyloid aggregates, present in so-called Lewy bodies. These, and the intermediate oligomeric aggregates present during the course of the aggregation process, are toxic to dopaminergic neurons and thus contribute to degeneration in Parkinson’s.
There are many NMR, X-Ray and Cryo-EM structures of αSyn monomer and fibrils in the PDB. (e.g. 1XQ8 [23], 2N0A [28] and 7C1D [29] . However, the picture emerging from these studies is complex. For example, the Cryo-EM studies have identified four distinct types of full length αSyn fibril to date, known as type 1a ‘rod’, type 1b ‘twister’, type 2a and type 2b polymorphs [30].
For the majority of these structural programmes the αSyn was expressed in and purified from E. coli with anion exchange chromatography often being included, due to the low 4.67 pI of αSyn. Stephens et al [31] provide an excellent review of the different methods that have been used to produce monomeric αSyn with comment on the impact of various steps (such as low pH) on its subsequent functionality.

Within the Protein Science and Structural Biology department at Sygnature Discovery we have been purifying various forms of αSyn and other fibril forming proteins for several years. The feedback that we have had is that the monomeric human wild type αSyn we make is of the highest quality with excellent batch to batch consistency and stability, that displays no self-seeding when used as a substrate within seed aggregation and RT-QuIC assays.
If this is of interest to your project please dont hesitate to get in touch with us to discuss your particular needs.
[1] L. Maroteaux, J. Campanelli, and R. Scheller, ‘Synuclein: a neuron-specific protein localized to the nucleus and presynaptic nerve terminal’, J. Neurosci., vol. 8, no. 8, pp. 2804–2815, Aug. 1988, doi: 10.1523/JNEUROSCI.08-08-02804.1988.
[2] K. Uéda et al., ‘Molecular cloning of cDNA encoding an unrecognized component of amyloid in Alzheimer disease.’, Proc. Natl. Acad. Sci., vol. 90, no. 23, pp. 11282–11286, Dec. 1993, doi: 10.1073/pnas.90.23.11282.
[3] J. Burré, ‘The Synaptic Function of α-Synuclein’, J. Parkinsons. Dis., vol. 5, no. 4, pp. 699–713, Oct. 2015, doi: 10.3233/JPD-150642.
[4] L. M. A. Oliveira et al., ‘Alpha-synuclein research: defining strategic moves in the battle against Parkinson’s disease’, npj Park. Dis., vol. 7, no. 1, p. 65, Dec. 2021, doi: 10.1038/s41531-021-00203-9.
[5] H. Grosso Jasutkar, S. E. Oh, and M. M. Mouradian, ‘Therapeutics in the Pipeline Targeting α -Synuclein for Parkinson’s Disease’, Pharmacol. Rev., vol. 74, no. 1, pp. 207–237, Jan. 2022, doi: 10.1124/pharmrev.120.000133.
[6] R. Jakes, M. G. Spillantini, and M. Goedert, ‘Identification of two distinct synucleins from human brain’, FEBS Lett., vol. 345, no. 1, pp. 27–32, May 1994, doi: 10.1016/0014-5793(94)00395-5.
[7] A. Iwai et al., ‘The precursor protein of non-Aβ component of Alzheimer’s disease amyloid is a presynaptic protein of the central nervous system’, Neuron, vol. 14, no. 2, pp. 467–475, Feb. 1995, doi: 10.1016/0896-6273(95)90302-X.
[8] M. Nakai et al., ‘Expression of α-synuclein, a presynaptic protein implicated in Parkinson’s disease, in erythropoietic lineage’, Biochem. Biophys. Res. Commun., vol. 358, no. 1, pp. 104–110, Jun. 2007, doi: 10.1016/j.bbrc.2007.04.108.
[9] S. Chandra, X. Chen, J. Rizo, R. Jahn, and T. C. Südhof, ‘A Broken α-Helix in Folded α-Synuclein’, J. Biol. Chem., vol. 278, no. 17, pp. 15313–15318, Apr. 2003, doi: 10.1074/jbc.M213128200.
[10] J. Varkey et al., ‘Membrane Curvature Induction and Tubulation Are Common Features of Synucleins and Apolipoproteins’, J. Biol. Chem., vol. 285, no. 42, pp. 32486–32493, Oct. 2010, doi: 10.1074/jbc.M110.139576.
[11] G. Fusco et al., ‘Direct observation of the three regions in α-synuclein that determine its membrane-bound behaviour’, Nat. Commun., vol. 5, no. 1, p. 3827, Sep. 2014, doi: 10.1038/ncomms4827.
[12] M. H. Polymeropoulos et al., ‘Mutation in the α-Synuclein Gene Identified in Families with Parkinson’s Disease’, Science (80-. )., vol. 276, no. 5321, pp. 2045–2047, Jun. 1997, doi: 10.1126/science.276.5321.2045.
[13] R. Krüger et al., ‘AlaSOPro mutation in the gene encoding α-synuclein in Parkinson’s disease’, Nat. Genet., vol. 18, no. 2, pp. 106–108, Feb. 1998, doi: 10.1038/ng0298-106.
[14] J. J. Zarranz et al., ‘The new mutation, E46K, of α-synuclein causes parkinson and Lewy body dementia’, Ann. Neurol., vol. 55, no. 2, pp. 164–173, Feb. 2004, doi: 10.1002/ana.10795.
[15] S. Appel-Cresswell et al., ‘Alpha-synuclein p.H50Q, a novel pathogenic mutation for Parkinson’s disease’, Mov. Disord., vol. 28, no. 6, pp. 811–813, Jun. 2013, doi: 10.1002/mds.25421.
[16] C. Proukakis et al., ‘A novel -synuclein missense mutation in Parkinson disease’, Neurology, vol. 80, no. 11, pp. 1062–1064, Mar. 2013, doi: 10.1212/WNL.0b013e31828727ba.
[17] S. Lesage et al., ‘G51D α-synuclein mutation causes a novel Parkinsonian-pyramidal syndrome’, Ann. Neurol., vol. 73, no. 4, pp. 459–471, Apr. 2013, doi: 10.1002/ana.23894.
[18] P. Pasanen et al., ‘A novel α-synuclein mutation A53E associated with atypical multiple system atrophy and Parkinson’s disease-type pathology’, Neurobiol. Aging, vol. 35, no. 9, pp. 2180.e1-2180.e5, Sep. 2014, doi: 10.1016/j.neurobiolaging.2014.03.024.
[19] H. Yoshino et al., ‘Homozygous alpha-synuclein p.A53V in familial Parkinson’s disease’, Neurobiol. Aging, vol. 57, pp. 248.e7-248.e12, Sep. 2017, doi: 10.1016/j.neurobiolaging.2017.05.022.
[20] H. Liu et al., ‘A Novel SNCA A30G Mutation Causes Familial Parkinsonʼs Disease’, Mov. Disord., vol. 36, no. 7, pp. 1624–1633, Jul. 2021, doi: 10.1002/mds.28534.
[21] R. Bussell and D. Eliezer, ‘A Structural and Functional Role for 11-mer Repeats in α-Synuclein and Other Exchangeable Lipid Binding Proteins’, J. Mol. Biol., vol. 329, no. 4, pp. 763–778, Jun. 2003, doi: 10.1016/S0022-2836(03)00520-5.
[22] W. S. Davidson, A. Jonas, D. F. Clayton, and J. M. George, ‘Stabilization of α-Synuclein Secondary Structure upon Binding to Synthetic Membranes’, J. Biol. Chem., vol. 273, no. 16, pp. 9443–9449, Apr. 1998, doi: 10.1074/jbc.273.16.9443.
[23] T. S. Ulmer, A. Bax, N. B. Cole, and R. L. Nussbaum, ‘Structure and Dynamics of Micelle-bound Human α-Synuclein’, J. Biol. Chem., vol. 280, no. 10, pp. 9595–9603, Mar. 2005, doi: 10.1074/jbc.M411805200.
[24] A. Oueslati, M. Fournier, and H. A. Lashuel, ‘Role of post-translational modifications in modulating the structure, function and toxicity of α-synuclein’, 2010, pp. 115–145.
[25] A. J. Trexler and E. Rhoades, ‘N-terminal acetylation is critical for forming α-helical oligomer of α-synuclein’, Protein Sci., vol. 21, no. 5, pp. 601–605, May 2012, doi: 10.1002/pro.2056.
[26] H. A. Lashuel, C. R. Overk, A. Oueslati, and E. Masliah, ‘The many faces of α-synuclein: from structure and toxicity to therapeutic target’, Nat. Rev. Neurosci., vol. 14, no. 1, pp. 38–48, Jan. 2013, doi: 10.1038/nrn3406.
[27] J. A. Steiner, E. Quansah, and P. Brundin, ‘The concept of alpha-synuclein as a prion-like protein: ten years after’, Cell Tissue Res., vol. 373, no. 1, pp. 161–173, Jul. 2018, doi: 10.1007/s00441-018-2814-1.
[28] M. D. Tuttle et al., ‘Solid-state NMR structure of a pathogenic fibril of full-length human α-synuclein’, Nat. Struct. Mol. Biol., vol. 23, no. 5, pp. 409–415, May 2016, doi: 10.1038/nsmb.3194.
[29] H. Long et al., ‘Wild-type α-synuclein inherits the structure and exacerbated neuropathology of E46K mutant fibril strain by cross-seeding’, Proc. Natl. Acad. Sci., vol. 118, no. 20, May 2021, doi: 10.1073/pnas.2012435118.
[30] R. M. Meade, D. P. Fairlie, and J. M. Mason, ‘Alpha-synuclein structure and Parkinson’s disease – lessons and emerging principles’, Mol. Neurodegener., vol. 14, no. 1, p. 29, Dec. 2019, doi: 10.1186/s13024-019-0329-1.
[31] A. D. Stephens, D. Matak-Vinkovic, A. Fernandez-Villegas, and G. S. Kaminski Schierle, ‘Purification of Recombinant α-synuclein: A Comparison of Commonly Used Protocols’, Biochemistry, vol. 59, no. 48, pp. 4563–4572, Dec. 2020, doi: 10.1021/acs.biochem.0c00725



Peak Proteins, NuChem Sciences, and SB Drug Discovery have now fully integrated with Sygnature Discovery.
